 فارسی
فارسی
 English
English


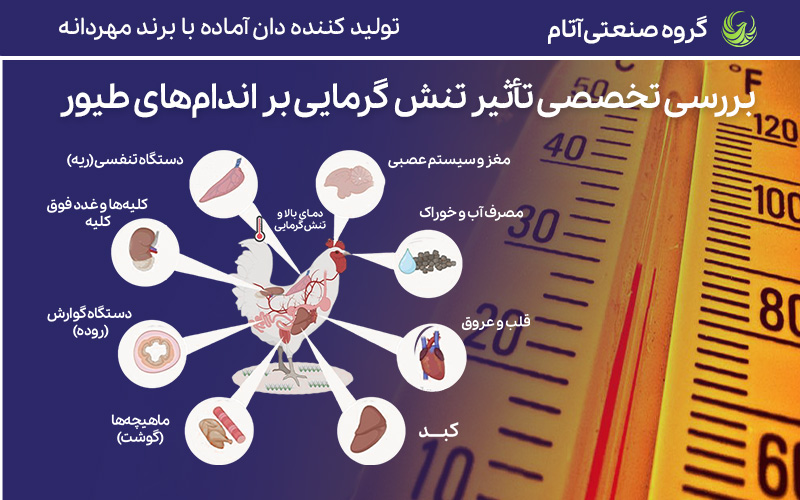
اثرات استرس گرمایی بر فیزیولوژی مرغها
جوجههای گوشتی امروزی که بر اثر حدود ۷۰ سال اصلاح ژنتیکی با رشد بسیار بالا، نسبت به سویههای با رشد آهستهتر، تحمل گرمایی کمتری دارند و آسیبپذیری بالاتری در برابر استرس گرمایی دارند. این مورد ناشی از نرخ متابولیک فوقالعاده بالا و رشد ناکافی سیستم قلبیعروقی و تنفسی است. در این رابطه گروهی از محققین اخیراً ثابت کردهاند که پاسخ فیزیولوژیک به استرس گرمایی در جوجههای سنگین نسبت به پرندگان سبکتر همان نژاد و سن، شدیدتر است و تحمل گرما در آنها کمتر است.
دیدگاههایی درباره فیزیولوژی و سلامت روده
علائم بالینی و پیامدهای استرس گرمایی چندوجهی است و طیف وسیعی از تغییرات را شامل میشود؛ از جمله تغییرات رفتاری (مانند بیحالی، کاهش مصرف خوراک و لهله زدن)، تغییرات متابولیک (مانند وضعیت کاتابولیک، تجمع چربی و کاهش رشد عضله)، بهم خوردن هموستاز عمومی (مثل آلکالوز، بههم خوردن تعادل هورمونی، نقص ایمنی، التهاب و استرس اکسیداتیو) و اختلال در عملکرد دستگاه گوارش (مانند اختلال در هضم و جذب، رودهآزردگی، نارسایی سد پارانشیمی و بهم خوردن توازن میکروبی روده یا دیسبیوز).
نقش تغذیه در مقابله با استرس گرمایی ثابتشده است. استراتژیهای تغذیهای که باعث افزایش بهرهبرداری از پروتئین و انرژی میشود و مداخلات تغذیهای که به بازگرداندن تعادل میکروبی روده کمک کند، به علت اهمیت بالای اثرات استرس گرمایی بر مصرف خوراک، متابولیسم مواد مغذی و سلامت روده، روزبهروز اهمیت بیشتری پیدا میکنند.
استرس گرمایی و رفتار پرنده
رفتارهای خنککننده (“Cooling behaviors”) بارزترین تغییرات رفتاری ناشی از استرس گرمایی در مرغها هستند. با بالا رفتن دمای محیط، اختلاف دمای بین سطح بدن و محیط کمتر میشود و اتلاف گرمای محسوس کاهش مییابد. بنابراین، مرغهایی که دچار هایپرترمی محیطی میشوند، با افزایش نرخ تنفس (تندنفسی/ لهله زدن)، سعی میکنند با تبخیر آب از دستگاه تنفسی، گرمای نهان بیشتری دفع کنند. اگرچه دفع گرمای محسوس به اختلاف دما وابسته است، اما رطوبت نسبی هم سقفی برای میزان تبخیر (و اتلاف گرمای نهان) ایجاد میکند. بنابراین، دمای بالا همراه با رطوبت بالا، به شدت توانایی دفع گرما را کاهش داده و اثرات زیانبار استرس گرمایی را تشدید میکند.
در شرایط استرس گرمایی طولانیمدت، لهله زدن به یک فاز آهستهتر و عمیقتر تبدیل میشود که به آن هایپرپنه گرمایی (thermal hyperpnea) گفته میشود. اگرچه لهله زدن، اتلاف گرمای نهان را افزایش میدهد، اما معایبی هم دارد؛ مهمترین آن، دهیدراسیون (کمآبی) است که باعث افزایش نیاز و مصرف آب میشود. همچنین لهله زدن منجر به دفع بیشتر CO₂، ایجاد هایپوکاپنی و در نهایت آلکالوز تنفسی میشود. آلکالوز تهدیدی برای صنعت تخممرغ است؛ چون باعث کاهش کلسیم یونیزه شده خون شده و معدنی شدن پوسته تخممرغ را مختل میکند. البته، آلکالوز ناشی از استرس گرمایی برای تولیدکنندگان جوجه گوشتی نیز تهدیدی جدی محسوب میشود.
مرغهای دچار استرس گرمایی اغلب بالهای خود را بلند میکنند تا قسمتهای بدون پر بدن را برای دفع گرمای محسوس بیشتر در معرض هوا قرار دهند. با وجود اینکه لهله زدن و بال بلند کردن برای تنظیم دمای بدن ضروری است، اما این فعالیتها انرژیبر هستند، و بخشی از کالری که باید صرف رشد و تولید شود، هدر میرود.
مرغهایی که در دماهای بالا نگهداری میشوند، کُند و بیحال شده، وقت بیشتری را به استراحت (مثلاً نشستن نزدیک به زمین) و زمان کمتری را به تغذیه و قدم زدن میگذرانند. این موضوع، مصرف خوراک و سلامت اسکلتی را تحت تأثیر منفی قرار میدهد. محدود کردن مصرف خوراک، مکانیزم محافظتی قدیمی برای کاهش گرمای تولیدی هنگام هضم، جذب و مصرف مواد مغذی است. عملکرد پایینتر مرغهای دچار استرس گرمایی همواره به کاهش مصرف خوراک نسبت داده شده است، اما مدلهای «تغذیه جفتسازیشده» نشان دادهاند که کاهش وزن بدن در جوجههای گوشتی در شرایط استرس گرمایی، به بیاشتهایی ربطی ندارد.
استرس گرمایی و متابولیسم چربی
اثر مستقیم استرس گرمایی بر فیزیولوژی، علاوه بر کاهش اشتها، نقش مهمی در اختلال عملکردی مرغها دارد. به طور پارادوکسیکال، حیوانات دچار استرس گرمایی با اینکه در حالت تعادل منفی انرژی و وضعیت کاتابولیک هستند، دچار محدودیت در بسیج چربی میشوند.
در واقع، مرغها و دامهای شیری نگهداری شده در محیطهای گرم، کاهش پیشرونده اسیدهای چرب آزاد غیر استریفیه (NEFA) را نشان میدهند که شاخص قابل اعتمادی برای بررسی متابولیسم چربی است و محدودیت استفاده از ذخایر چربی را نشان میدهد.
تحقیقات متعدد نشان دادهاند که مرغهای دچار استرس گرمایی، چربی احشایی (شکمی)، زیرجلدی و میانعضلهای بیشتری ذخیره میکنند. تجمع بیشتر چربی در نقاط سطحی بدن، خود باعث میشود پرنده نتواند گرما را دفع کند و خطر هایپرترمی شدید را افزایش میدهد.
شایان ذکر است که تغییرات متابولیسم چربی فقط به کاهش استفاده از ذخایر چربی محدود نمیشود؛ چون مرغهای دچار استرس گرمایی همچنین افزایش بیان پروتئینهای لیپوژنز جدید در کبد و تجمع چربی در این اندام را نشان میدهند.
استرس گرمایی و متابولیسم پروتئین عضلات اسکلتی
علاوه بر افزایش مقدار چربی، ثابت شده که استرس گرمایی ترکیب لاشه جوجههای گوشتی را با کاهش نسبت بافت بدون چربی، بهخصوص بازدهی سینه، تغییر میدهد. نخستین بررسیهای مولکولی نشان دادند که کاهش رسوب پروتئین عضله ناشی از استرس گرمایی، عمدتاً به کاهش سنتز پروتئین مربوط است، نه تجزیه شدیدتر پروتئین. با این حال، مطالعات نشان داد که علت کاهش تجمع بافت بدون چربی میتواند بسته به نوع عضله متفاوت باشد: در سینه کاهش سنتز پروتئین و در ران، افزایش تجزیه پروتئین دیده میشود. همچنین، آنها اختلال در سنتز پروتئین را با کاهش بیان عوامل رشد شبه انسولینی نوع ۱ (IGF-1)، PI3K و p70S6 کیناز (S6K)، و افزایش تجزیه پروتئین را با افزایش بیان MAFbx یا آتروژین-۱ مرتبط دانستند. دیگر دانشمندان نیز این تغییرات در بیان S6K و MAFbx ناشی از استرس گرمایی را تأیید کردند. S6K در کنترل سنتز پروتئین و رشد عضله در مرغها حیاتی است.
جالب است بدانید گروهی از محققین اعلام کردند مسیر سیگنالدهی S6K در پاسخ به محرک آنابولیک در جوجههای گوشتی دچار استرس گرمایی واکنش محدودی دارد. در یک آزمایش افزایش اسید اوریک، اوره و اسیدهای آمینه پروتئینی خون را ثبت کردند، با وجود کاهش محسوس مصرف خوراک و بازدهی سینه که همراه با کاهش گلوکز و NEFA بود، محققین فرض کردند که جوجههای در معرض گرما برای جبران ناتوانی در استخراج انرژی از چربی ذخیرهای، به سراغ ذخایر پروتئینی عضلات اسکلتی، بهویژه سینه، میروند. در این راستا، سطح پلاسمایی کراتین، ۳-متیل هیستیدین و اوره به عنوان نشانگر بررسی تجزیه پروتئین عضلانی ناشی از استرس گرمایی بهکار میرود.
این فرضیه نیز با تحقیقات دیگر دانشمندان حمایت شد: آنها یافتند که استرس گرمایی اسیدآمینههای گلوکوژنیک پلاسمایی را کاهش میدهد، جذب اسیدآمینه کبد و پتانسیل گلوکوژنیک آن را افزایش میدهد و فعالیت ترانسآمینازهای کبدی را برای دیآمینهسازی اسیدآمینهها (جهت پیشسازسازی گلوکونئوژنز) تقویت میکند. همچنین اگرچه در جوجههای دچار استرس گرمایی سطوح اسیدآمینههای پیشساز گلوکز و اسیدآمینه آزاد عضله سینه کاهش مییابد، اما همزمان اسیدآمینههای سازنده پروتئین در خون افزایش یافته و مصرف خوراک بهشدت کاهش مییابد.
استرس گرمایی و سطوح هورمونی
مرغهایی که دچار استرس گرمایی میشوند، تغییرات هورمونی مشابهی با پستانداران دارند. استرس گرمایی محور هیپوتالاموس-هیپوفیز-آدرنال را فعال و باعث افزایش قابلتوجه گلوکوکورتیکوئیدها، بهویژه کورتیکوسترون، در خون میشود.
در مرغها، سطوح بالای کورتیکوسترون باعث کاهش رشد، القای پروتئولیز (تجزیه پروتئین) و مهار سنتز پروتئین در عضلات اسکلتی و نیز افزایش ذخیره چربی میشود، که همگی از پیامدهای کلاسیک استرس گرمایی هستند. فرض بر این است که کورتیکوسترون با تغییرات مذکور در بیان S6K و MAFbx بر متابولیسم پروتئین عضله اثر منفی میگذارد و همزمان با افزایش بیان سنتتاز اسید چرب (FASN) در سلولهای کبدی و چربی، اثر لیپوژنیک دارد. با این حال، پژوهش جدیدی نشان داده که تیمار سلولهای عضلانی مرغ در شرایط آزمایشگاهی با کورتیکوسترون، تجزیه پروتئین و بیان MAFbx را نسبت به شرایط استرس گرمایی بهتنهایی افزایش نمیدهد، بنابراین برای روشن شدن نقش کورتیکوسترون در تغییرات متابولیسم پروتئین ناشی از استرس گرمایی به بررسی بیشتری نیاز است.
علاوهبر این، چون افزایش کورتیکوسترون منجر به تضعیف عملکرد ایمنی میشود، مرغهای دچار استرس گرمایی ایمنی ضعیفتری داشته و مستعد ابتلا به بیماریهای عفونی هستند. اختلالات جدی ایمنی نظیر کاهش تولید ایمونوگلوبولین در برابر آنتیژن نمونه، آتروفی و اختلال عملکرد بافتهای لنفاوی اولیه و ثانویه همراه با کاهش لنفوسیت در جوجههای گوشتی استرس دیده ثبت شده است. محققین، کورتیکوسترون بالا را از دلایل محتمل این نابسامانیهای ایمنی دانستند.
کاهش سطح خونی ترییدوتیرونین (T3) و تیروکسین (T4) هم در مرغهای تخمگذار و هم در جوجههای گوشتی مشاهده شده است. این تغییرات، که احتمالاً ناشی از کاهش اندازه و فعالیت تیروئید است، در گاو شیری دچار استرس گرمایی نیز ثبت شده است. معمولاً فرض میشود پاسخ تیروئید به دمای بالای محیط، نوعی سازوکار تطبیقی است که به حیوانات امکان میدهد متابولیسم پایه و تولید گرما را پایین بیاورند و از بیشگرم شدن پیشگیری کنند. این وضعیت شبیه کمکاری تیروئید، میتواند تا حدی افسردگی رشد، افزایش چربی لاشه و کاهش تولید تخممرغ و کیفیت پوسته را در زمان استرس گرمایی توجیه کند.
تأثیر استرس گرمایی بر سلامت روده مرغها
سلامت روده باید به شکل جامع دیده شود و سه عامل اصلی که به طور همافزا بر آن اثر دارند را در نظر گرفت: اپیتلیوم (سطح مخاطی) دستگاه گوارش، سیستم ایمنی اختصاصی روده، و میکروبیوتای رودهای. دستگاه گوارش، به عنوان بزرگترین سطح در تماس با محیط، مرتباً توسط عوامل مضر مختلفی تهدید میشود. مانند ترکیبات مضر موجود در خوراک و میکروارگانیسمهای بیماریزا.
وظایف دستگاه گوارش با هم متعارض است: از یک سو باید نهایت جذب مواد مغذی را انجام دهد، و از سوی دیگر باید انواع محرکهای آنتیژنی را شناسایی کند و در عین حال، میکروبیوتای مفید ساکن را تحمل نماید. بنابراین، حفظ یکپارچگی و عملکرد مناسب ساختاری و فیزیولوژیکی این دستگاه برای سلامتی و کارایی مرغها حیاتی است.
استرس گرمایی و اپیتلیوم دستگاه گوارش
اپیتلیوم گوارشی که به صورت یک لایه سلولی مفرد آرایش یافته، نقش فعالی در سیستم ایمنی مجتمع روده دارد؛ این لایه، سدی است که توسط پروتئینهای اتصال محکم (Tight Junction – TJ) تقویت شده، موسیلاژ (موکوس) و پپتیدها و پروتئینهای ضد میکروبی نیز ترشح میکند و از گیرندههای شناسایی الگو (PRR) برای سازماندهی پاسخ ایمنی موضعی استفاده میکند.
اتصالات محکم (TJ)، که بالاترین بخش مجموعه اتصالات نوک راسی سلول هستند، فضای بین سلولهای ستونی اپیتلیال را میبندند. این اتصالات شامل پروتئینهای ترانسممبران (کلودینها و اوکلودین) و پروتئینهای اسکلتی-تکیهگاهی مانند(ZO) هستند. پروتئینهای ZO یک طرف خود را به کلودین/اوکلودین و طرف دیگر را به حلقه آکتومایوزین متصل میکنند، پس واسطهای بین TJ ترانسممبران و اسکلت سلولی هستند.
وظیفه اصلی TJها کنترل مسیر عبور بین سلولی است. این مسیر برخلاف انتقال فعال سلولی، انتقال غیرفعال (انتشار ترانساپیتلیال) را از طریق دو مسیر اصلی امکانپذیر میکند: مسیر منفذی (تحت کنترل کلودینها؛ عبور محدود ملکولهای باردار و بزرگتر از ۴ آنگستروم) و مسیر نشتی(تحت کنترل اوکلودین و ZO؛ عبور ملکولهای بزرگتر از جمله LPS باکتریها).
در شرایط استرس گرمایی، بدن مرغ برای دفع مؤثرتر گرما، خون را از بافتهای داخلی (شکمی) به محیط (پوست و اندامها) میفرستد، که به قیمت کاهش خونرسانی به دستگاه گوارش تمام میشود. این کاهش خونرسانی (هیپوپرفیوژن) باعث رسوب کمتر مواد مغذی و اکسیژن به روده شده و به لایه مخاطی آسیب میزند.
از سوی دیگر، دستگاه گوارش از نظر نیاز انرژی و پروتئین بسیار پرمصرف است و آسیب تغذیهای باعث کاهش بازسازی سلولی و تضعیف سد روده میشود.
همزمان، کمبود اکسیژن (هیپوکسی) باعث تغییر عمیق در متابولیسم سلول و افزایش تولید گونههای فعال اکسیژن و نیتروژن (ROS ,RNS) میشود.
خود هایپرترمی هم تولید ROS و RNS را بالا میبرد و سیستمهای آنتیاکسیدانی سلولی را تضعیف میکند. دمای بالا مستقیماً استرس اکسیداتیو ایجاد میکند، زنجیره تنفسی میتوکندری را مختل و سبب تولید بیش از حد ROS و آسیب اکسیداتیو میشود.
در عضلات اسکلتی علیالخصوص سینه (سینه مرغ)، افزایش پتانسیل غشای میتوکندری، تولید بالای سوپراکسید و ROS و همچنین افزایش مالوندیآلدهید (MDA) به عنوان شاخص پراکسیداسیون لیپیدی دیده شده است.
در مورد روده، استرس اکسیداتیو سد پاراسلولار تحت کنترل TJ را بیثبات و نفوذپذیری روده را زیاد میکند.
کیناز زنجیره سبک میوزین (MLCK) نیز در این فرایند نقش دارد؛ زیرا انقباض حلقه آکتومایوزین را کنترل و غیرمستقیم مسیر پاراسلولار را تنظیم میکند. استرس اکسیداتیو میتواند انقباض آکتومایوزین و جابهجایی و کاهش بیان پروتئین ZO را القا کند و سد پاراسلولار را بیشتر تضعیف نماید. با افزایش کورتیکوسترون در استرس گرمایی نیز سد مخاطی تضعیف میشود.
برای سنجش یکپارچگی سد پاراسلولار معمولاً مقاومت الکتریکی تراپیتیلیالی (TEER) و عبور پروبهای نشانهدار از مخاط به سروز (مانند فلورسئین ایزوتیوسیانات-دکستران) بررسی میشود. سد سالم TEER بالا و عبور پروب کم دارد، اما استرس گرمایی TEER را پایین میآورد و عبور نشانگر را افزایش میدهد؛ یعنی روده «سوراخدار» شده و مواد مضر به راحتی عبور میکنند. این حالت در مدلهای مختلف حیوانی و جوجه گوشتی اثبات شده است.
افزایش نفوذپذیری روده باعث عبور الگوهای ملکولی مرتبط با میکروب (PAMP) – عمدتاً LPS باکتریهای گرممنفی – به لایههای زیرمخاطی میشود. این LPSها توسط گیرندههای TLR4–MD-2 در سطح قاعدهای سلولهای روده شناسایی شده و سیگنالگذاری برای بیان سایتوکاینهای التهابی را فعال میکنند.
این سایتوکاینها TNF-α)، IL-1β، IFN-γ ) سد روده را تضعیف، باعث نشت LPS و پیشبرد چرخه التهاب و زخم میشوند و موجب انقباض آکتومایوزین و جابهجایی اوکلودین و جداشدن TJها خواهند شد.
این التهاب موضعی موجب کوتاه شدن عمر سلولهای رودهای، هیپرپلازی کریپت و آتروفی پرزهای روده میشود. این اختلالات مورفولوژیک (به همراه افزایش آپوپتوز و کاهش تکثیر سلولی) اخیراً در جوجه گوشتیهای دچار گرما دیده شده است.
در مطالعهای نشان داده شد که آسیب ساختاری و افزایش نفوذپذیری روده ناشی از اثر مستقیم استرس گرمایی است، نه صرفاً کاهش مصرف خوراک. نتیجه این آسیبها؛ سوءجذب مواد مغذی و صرف انرژی بالا برای پاسخ ایمنی روده است که عملکرد مرغ را به شدت کاهش میدهد و میتواند زمینهساز سایر مشکلات سلامتی شود.
افزون بر ایجاد التهاب موضعی، LPSهای عبوری میتوانند وارد گردش پورتال شده به کبد رسیده و آن را درگیر کنند. اگر مقدار LPS از توان سمزدایی کبد بیشتر شود، وارد جریان خون عمومی میشود و سبب اندوتوکسمی، التهاب سیستمیک و اختلال عملکرد سایر بافتها میگردد. در حالت شدید، اندوتوکسمی به نارسایی چندعضوی و شوک سپتیک کشنده منتهی خواهد شد.
اختلالات در انتقال سلولی نیز از دلایل نفوذپذیری بالای روده در استرس گرمایی است؛ چون از دست رفتن یکپارچگی اپیتلیوم میتواند به آسیب سلولی و بازشدن مسیرهای مستقل از TJ بیانجامد. در این شرایط، باکتریهای رودهای راحتتر وارد کبد یا بافتها و اندامهای دیگر میشوند، که به این پدیده «ترانسلوکاسیون باکتریایی» میگویند. برای مثال، در جوجههای گوشتی دچار استرس گرمایی، تهاجم سالمونلا به کبد بیشتر است. این اتفاق میتواند مقدمه مشکلات خارج رودهای از جمله آسیب کبد و کندرو نکروز با استئومیلیت (BCO) باشد.
استرس گرمایی و میکروبیوتای دستگاه گوارش
اصطلاحات "میکروبیوتا" و "میکروبیوم" اغلب به جای هم به کار میروند، اما در واقع تفاوتهایی دارند. میکروبیوتا به مجموعه میکروارگانیسمهایی اشاره دارد که در یک محیط خاص (مثلاً بخشی از بدن انسان یا حیوان) زندگی میکنند، ولی میکروبیوم هم متاژنوم میکروبیوتا (یعنی مجموعه ژنهای میکروبها) و هم محیط اطراف آن را شامل میشود.
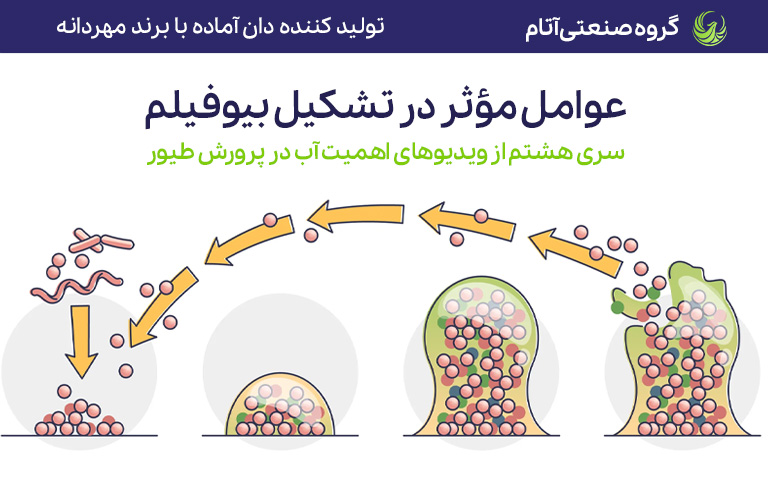
دستگاه گوارش مرغها جمعیت میکروبی بسیار پیچیدهای دارد که شامل باکتریها، آرکئا، پروتوزوآ، قارچها و ویروسهاست. میکروبیوتای دستگاه گوارش عملاً ژنوم میزبان را توسعه داده و به طور قابل توجهی فیزیولوژی پرنده را تحت تأثیر قرار میدهد و تقریباً نقش یک اندام مکمل یا حتی اندام فراموششده را ایفا میکند.
دیدگاه غالب امروزی این است که میکروبیوتای گوارش در برنامهریزی و تنظیم هر دو سیستم ایمنی موضعی و سیستم ایمنی عمومی بدن، از جمله در طیور نقش بسیار مهمی دارد. این نظریه با استفاده از مدلهای گنوتوبیوتیک نیز تأیید شده است؛ به طوری که گزارش شده است موشهای بدون میکروب و مرغهای بدون میکروب دچار نقصها و کمبودهای شدید رشدی و اختلال در سیستم ایمنی دستگاه گوارش میشوند. افزون بر نقش ایمنی، میکروبیوتای گوارش به شکل قابل توجهی رشد، مورفولوژی و کارکرد روده در مرغ را تحت تأثیر قرار میدهد.
عوامل فراوانی از جمله عوامل مربوط به میزبان و محیط، ساختار میکروبیوتای روده را تغییر میدهند. شواهد متعددی نشان داده که دمای بالای محیط میتواند به شدت ساختار جمعیت میکروبی روده را تغییر دهد. مطالعات متعدد ثابت کردهاند که استرس گرمایی (HS) باعث اختلال در میکروبیوتای رودهای در موش، طیور و گاو شیری شده و تعادل آن را مختل (دیسبیوزیس) میکند.
دیسبیوزیس (یا دیسباکتریوز) به تغییر ترکیب میکروبیوتای روده گفته میشود که معمولاً با ازدیاد میکروبهای مضر یا کاهش جمعیت باکتریهای مفید همراه است و تعادل شکننده بین میزبان و میکروبیوتای او را دستخوش ضعف میکند.
وضعیت دیسبیوزیس اغلب با کاهش هضم مواد مغذی، از دست دادن کارکرد سد روده و التهاب رودهای همراه است، در حالی که یوبیوزیس، یعنی تعادل سالم میکروبی، میتواند سلامت، بهرهوری و توان پرندگان برای مقابله با استرسهای محیطی را افزایش دهد.
اگرچه امروزه روشهای پیشرفته آنالیز برای بررسی میکروبیوتای روده وجود دارد، اما تغییرات ساختاری، ترکیبی و عملکردی میکروبیوتای روده ای مرغهای تحت استرس گرمایی هنوز به طور کامل روشن نشده است. با این حال، تغییر شکل روده، تغییر مقدار و ترکیب موکوس و نقاط اتصال، همراه با تجمع ترکیبات خوراکی هضمنشده/کمهضم، همگی دلایلی محتمل برای دیسبیوزیس ناشی از استرس گرمایی هستند.
میکروبیوتای همزیست میتواند از کلونیزاسیون و رشد میکروارگانیسمهای بیگانه و بیماریزا در دستگاه گوارش جلوگیری کند. این مکانیزم حفاظتی به عنوان حذف رقابتی (Competitive Exclusion) یا مفهوم نورمی شناخته میشود و نخستین بار در جوجههای تازه از تخم درآمده نشان داده شد که اگر با سوسپانسیون محتویات چینهدان و روده مرغهای بالغ سالم تغذیه شوند، مقاومت بالایی به سالمونلا پیدا میکنند.
همچنین، باکتریهای مفید قادرند در رقابت برای مواد مغذی و تولید مواد بازدارنده و کشنده میکروب (مانند اسیدهای آلی، باکتریوسینها) پاتوژنها را کنترل کنند.
اما، میکروبیوتای مختلآمیز (غیرطبیعی) در مرغهای دچار استرس گرمایی با افزایش پذیرش به کلونیزاسیون رودهای سالمونلا انتریتیدیس همراه بوده است. نشان دادند که استرس گرمایی میتواند رشد کلستریدیوم پرفرنجنس در روده مرغ را افزایش داده و عامل مستعدکننده بیماری انتریت نکروتیک در گلههایی شود که در شرایط گرم پرورش مییابند. کلستریدیوم پرفرنجنس میتواند انتروتوکسینهایی ترشح کند که همراه با سایر عوامل مخرب باکتریایی، اتصالات محکم و عملکرد سد روده را مختل مینماید.
در مجموع، دیسبیوزیس، اختلال سد روده و التهاب مخاط با هم مرتبط بوده و اثرات منفی استرس گرمایی بر سلامت روده، فیزیولوژی و کارایی مرغها را تشدید میکنند.
نتیجهگیری
طبق مقالات مروری موجود، استرس گرمایی بازهی وسیعی از اثرات مخرب را به ویژه در مرغهای نژادهای پرتولید امروزی ایجاد میکند . نخست، استرس گرمایی هموستاز ایمنی، تعادل هورمونی، و وضعیت التهابی و اکسیداتیو را بهطور منفی متأثر میکند. تحقیقات بیشتر بر این تغییرات فیزیولوژیک و ارتباطشان میتواند به توسعه راهکارهای چندجانبه برای مقابله بهتر مرغها با این استرس کمک کند.
دوم، استرس گرمایی باعث کاتابولیسم بافتی و تغییرات عمده در متابولیسم پروتئین و چربی میشود. شواهد زیادی هست که نشان میدهد استرس گرمایی از طریق هم مهار سنتز پروتئین و هم تحریک تجزیه پروتئین، رسوب بافت عضلانی مرغ را مختل میکند، اما هنوز نیاز به تحقیقات بیشتر برای روشنشدن دلایل کاهش آزادسازی چربی در این شرایط است.
در نهایت، دمای بالا عملاً یک «استرسور دیسبیوزیسزا» محسوب میشود که سبب اختلال عملکرد روده و بهمریختگی رابطه میزبان و میکروبیوتا میگردد. بنابراین تقویت سد روده، بازیابی فرایندهای هضمی و جذبی، ایجاد تعادل در میکروبیوتای روده و کاهش التهاب و استرس اکسیداتیو از اقدامات ضروری برای افزایش تحمل و تابآوری مرغها در برابر استرس گرمایی است.
در مجموع، بازگرداندن نظم هموستاتیک و متابولیک مختلشده توسط استرس گرمایی و ایجاد حفاظت گوارشی راهبردهایی امیدبخش در مقابله با این تهدید رو به رشد برای پایداری صنعت طیور به شمار میروند.






































































024532531.jpg?v=3Me6NG2_nIZ8WXVravM98a_fqZqLFWFs2xiJ9PUZq4A)


















































005501674.jpg?v=Tf8WTPxBKykjrdsOHwFqWCuAnE5XTw8TMCzymNi5ch0)


002226069.jpg?v=-9B5a0CcDYIVH1NSveV6FTmGe2r-kE3pbZhcGikxMKU)









005240285.jpg?v=pa7qIxfeulTU1ZxrGmmvOYjPl8H33JSqprB8FsfmDck)
003421766.jpg?v=Qkjs3wy10ezXHrlmr4Add3ksNJdNok5OS8LvvTUG8BU)
001716576.jpg?v=3QBhhkvX4x6nsPxUljY0c_2vOG_Qooasb9umgZ0Zy0M)